항체 소개
항체 소개
항체 (Antibody) 는 면역글로불린 (Ig) 이라고도 불리며, 항원 자극에 반응하여 면역 체계, 특히 B 림프구에서 생성되는 고도로 특이적인 당단백질류입니다. 이들은 적응성 체액 면역의 핵심 작용 분자로서 병원 감염으로부터 방어하고 비정상 세포를 제거하며 체내 항상성을 유지하는 데 대체할 수 없는 역할을 수행합니다.
I. 기본 구조 및 기능적 도메인
항체 분자의 기본 구조는 고전적인 "Y" 형의 4 가닥 펩타이드 사슬 대칭 구조로 특징지어집니다:
-두 개의 동일한 중쇄 (Heavy Chain, H chain): 분자량이크게 (약 50-75 kDa), 중쇄는 항체의 클래스와 서브클래스를 결정합니다.
-두 개의 동일한 경쇄 (Light Chain, L chain): 분자량이 작으며 (약 25 kDa) 경쇄는 카파 (κ) 와 람다 (λ) 유형으로 나뉩니다.
-연간 이황화 결합: 이 결합은 중쇄와 경쇄 (H-L) 를 연결하며, 두 개의 중쇄 (H-H) 도 연결하여 분자 구조의 안정성을 유지합니다.
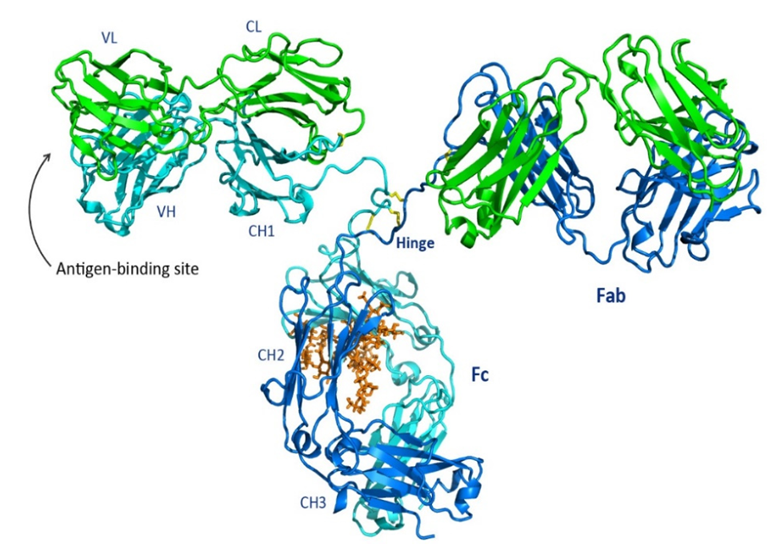
항체 분자는 프로테아제 (파파인, 펩신 등) 에 의해 특징적인 조각으로 가수분해될 수 있습니다:
-Fab 조각 (Fragment antigen-binding): 하나의 완전한 경쇄와 중쇄의 N-단부 (VH 와 CH1 도메인) 로 구성됩니다. 각 Fab 조각은 하나 이상의 항원 결합 부위를 가지며, 이는 항체의 특이성을 결정합니다. 이 부위는 경쇄와 중쇄의 가변 영역 (Variable Region, V region) 으로 형성되어 높은 변이도를 가진 아미노산 서열을 통해 항원에 직접 접촉하고 결합하는 상보적 결정 영역 (CDRs) 을 생성합니다.
-Fc 조각 (Fragment crystallizable): 두 개의 중쇄 C-단부 (IgG, IgA, IgD 의 CH2 와 CH3 도메인) 로 구성됩니다. Fc 조각은 항원과 결합하지는 않지만 항체의 작용 기능 (예:)
-보결합계 활성화 (고전적 경로를 통해).
-Fc 수용체를 발현하는 세포 (대식세포, NK 세포, 중성구, 호산구 등) 에 결합하여 옵소닌 포식작용, 항체 매개 세포 독성 (ADCC), 1 형 과민반응을 매개합니다.
-태반을 통과하거나 (IgG) 점막 표면으로 운반됩니다 (IgA).
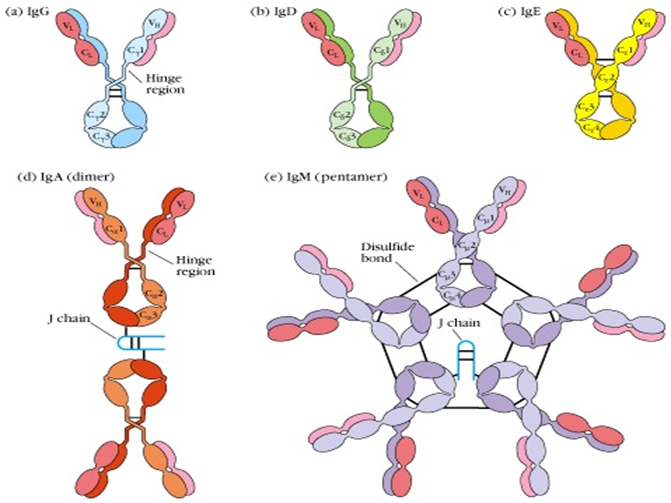
II. 면역글로불린의 종류와 서브클래스
중쇄의 상수 영역 (Constant Region, C region) 의 항원성과 구조에 따라 포유류 항체는 5 가지 주요 클래스 (isotype) 로 나뉩니다:
1. IgG: 혈장 내 가장 풍부하며 (70-75%), 분자량 약 150 kDa 입니다. 이는 2 차 면역 반응의 주요 항체로서 강력한 독소 및 바이러스 중화 능력을 가지며, 보결합을 활성화하고 옵소닌화를 매개합니다. 또한 태반을 통과하여 신생아에게 수동 면역을 제공하는 유일한 항체입니다. 인간 IgG 는 중쇄의 미세한 차이に基づ어 4 개의 서브클래스 (IgG1, IgG2, IgG3, IgG4) 로 더 나뉩니다.
2. IgM: 주로 펜타머 형태 (약 900 kDa) 로 존재하며, 1 차 면역 반응에서 처음 생성되는 항체 ("선구抗体") 입니다. 다수의 Fc 섹먼트를 가지므로 고전적 보결합 경로를 강력하게 활성화하여 초기 방어, 병원제 제거 및 면역복합체의 형성에 중요한 역할을 합니다. 단량형 IgM 은 주로 B 세포 막 표면에서 발현되어 B 세포 수용체 (BCR) 로 기능합니다.
3. IgA: 혈장 내에서는 주로 단량형이지만, 점막 관련 림프 조직 (MALT) 에서는 이이머나 트리머 형태로 형성되어 분비 성분을 결합하여 분비성 IgA (sIgA) 를 구성합니다. sIgA 는 점막 면역의 주요 장벽으로 타액, 눈물, 호흡기 및 소화액, 산유물에 존재하며 병원체가 점막 표면에 정착하고 침투하는 것을 방지합니다.
4. IgD: 단량형 형태 (약 180 kDa) 로 매우 낮은 혈장 농도로 존재합니다. 주요 기능은 성숙한 B 세포 표면의 항원 수용체 (BCR) 의 일부로서 작용하여 B 세포 활성화, 분화 및 면역 허용 유지를 관여합니다.
5. IgE: 단량형 형태 (약 190 kDa) 로 혈장 내 농도가 가장 낮습니다. 이의 Fc 섹먼트는 호산구와 기호구의 표면에 존재하는 고친화도 FcεRI 수용체에 결합합니다. 해당 항원 (알레르겐) 에 결합하면 이 세포들의 탈립질화를 유발하여 히스타민 및 기타 생리활성 매개체를 방출하고, 1 형 과민반응 (과민 반응) 및 항기생충 면역성을 매개합니다.
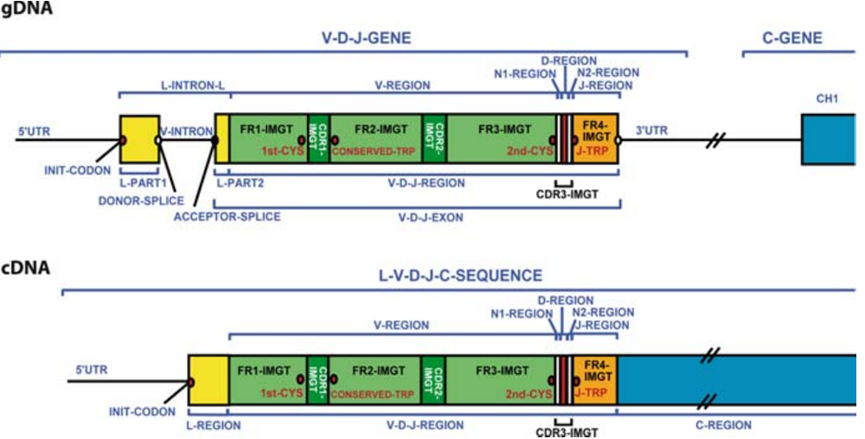
III. 항체 다양성 생성 메커니즘
항체 레퍼토리는 거의 무한한 다양성을 가지며 자연의 거의 모든 항원을 인식할 수 있습니다. 이 다양성은 다음과 같은 것에서 비롯됩니다:
1. 조개선 유전자 단편 재배열에 의한 조합적 다양성: V(D)J 유전자 재배열 (중쇄: VH, DH, JH 섹먼트; 경쇄: VL, JL 섹먼트) 은 다양한 V-영역 유전자 조합을 생성합니다.
2. 접합부 다양성: V-D, D-J, V-J 접합부에서 불완전한 뉴클레오타이드 절제 및 첨가 (P-뉴클레오타이드, N-뉴클레오타이드) 로 서열 변이가 발생합니다.
3. 체세포 초변이: 거대 중심에서 활성화된 B 세포는 V-영역 유전자에 고빈도 점 돌연변이를 겪어, 높은 친화도를 가진 항체의 항원 유도 선택을 통해 친화성 성숙 (affinity maturation) 을 달성합니다.
4. 중쇄와 경쇄의 무작위 결합: 다른 중쇄와 경쇄의 무작위 조합은 다양성을 더욱 증폭시킵니다.
5. 클래스 전환 재조합 (CSR): 사이토카인 및 기타 신호에 의해 유도되어 B 세포는 분비하는 항체의 등소형 (isotype) 을 변경할 수 있습니다 (IgM/IgD 에서 IgG, IgA 또는 IgE 로). 이는 항원 특이성 (V 영역은 변하지 않음) 을 유지하면서 다른 작용 기능을 획득하여 다양한 면역 요구에 부응합니다.
IV. 항체의 핵심 기능
1. 항원의 특이적 인식 및 결합: 이는 항체의 가장 기본적인 기능입니다. V-영역 CDR 을 통해 항체들은 항원 (예: 병원 표면 단백질, 독소, 외래 거대분자) 의 특정 에피토프에 정확하게 인식하고 결합합니다.
2. 중화작용: 항체들은 병원체 (바이러스, 박테리아 독소 등) 의 핵심 부위에 결합하여 숙주 세포 수용체와 상호작용하거나 세포 내로 침투하는 것을 방지함으로써 감염력이나 독성을 중화합니다.
3. 옵소닌화: 항체의 Fab 섹먼트 (주로 IgG) 는 입자성 항원 (예: 병원체) 에 결합하고, Fc 섹먼트는 포식 세포 (대식세포, 중성구 등) 의 FcγR 에 결합하여 이 세포들의 포식 효율을 현저히 증진시킵니다.
4. 보결합 활성화: 항원에 결합한 항체 (주로 IgM 과 IgG1/IgG3) 는 면역복합체를 형성하여 보결합계의 고전적 경로를 효과적으로 활성화합니다. 여기에서 다음과 같은 다양한 효과가 발생하며: 막 공격 복합체 (MAC) 를 형성하여 병원체나 세포를 용해시키고, 옵소닌인 (C3b) 을 생성하여 포식작용을 증진시키며,화학유인 인자를 (C3a, C5a) 생성하여 염증성 세포를 모집합니다.
5. 항체 매개 세포 독성 (ADCC): 항체의 Fab 섹먼트 (주로 IgG) 는 표적 세포 (바이러스 감염 세포 또는 종양 세포 등) 의 항원에 결합하고, Fc 섹먼트는 효과기 세포 (예: NK 세포) 의 FcγRIII (CD16) 에 결합하여 이 효과기 세포를 활성화해 퍼포린, 그랜자임 및 기타 세포 독성 분자를 방출시켜 표적 세포를 살해합니다.
6. 과민반응 매개: IgE 는 호산구/기호구의 FcεRI 에 결합하고, 알레르겐 교차 연결 시 탈립질화와 1 형 과민반응을 유발합니다. 일부 IgG/IgM 은 또한 2 형 (세포 독성) 과 3 형 (면역복합체 매개) 과민반응에도 관여할 수 있습니다.
7. 면역 조절: 항체는 이디오타 네트워크를 통해 면역 반응을 조절합니다. B 세포의 FcγRIIB 에 결합하는 Fc 섹먼트는 억제 신호를 전달하여 B 세포 활성을 낮춥니다. 항체 피드백 억제도 체액성 면역을 조절하는 중요한 메커니즘입니다.
항체는 면역 체계의 정교하고 강력한 무기입니다. 그들의 기발한 Y 형 구조, 특히 높은 변이도를 가진 항원 결합 부 (Fab) 와 상수 작용 영역 (Fc) 은 거의 무한한 다양한 항원을 인식하고 다각적인 면역 기능을 수행하는 능력을 부여합니다. 연구 도구로서 이들은 단백질 정제, 세포 또는 조직 위치 확인 연구 (WB, IHC, IF 등) 및 기능성 연구에서 결정적인 역할을 합니다.
EnkiLife 는 다양한 연구 수요를 충족시키기 위해 고품질 항체를 광범위하게 제공합니다. 항체 카탈로그 검색.
단클로날 항체 기술의 성숙과 항체 공학의 급속한 발전에 따라 항체는 기초 면역학적 연구 대상에서 현대 의학 진단, 예방 및 치료의 핵심으로 진화했습니다. 이들은 암 및 자가면역 질환 등 주요 질병 분야에서 혁신적인 치료 효과를 입증하며 생물 의학 혁신의 최전방을 이끌고 있습니다.
 | Dylan Z Dylan Z 는 EnkiLife 의 단백질 및 항체 전문가로 단백질 발현 시스템과 항체 준비 기술에 능통합니다. 그는 기술에서 완벽을 추구하며 사용자에게 안정적이고 사용하기 쉬운 제품을 개발하는 데 헌신합니다. |